Wang, C., & Kuzyakov, Y. (2024). Mechanisms and implications of bacterial-fungal competition for soil resources. The ISME journal, 18(1), wrae073.
https://doi.org/10.1093/ismejo/wrae073
转发:根际微生态, 2025.06.26
细菌和真菌之间的复杂相互作用决定了土壤中微生物群落的结构、组成和功能,并调控碳和营养物质的流动,对于理解生物地球化学循环至关重要。在各种相互作用中,资源竞争是决定这两大微生物群体在土壤中适应和生态位分化的主要因素。这是因为微生物生长所需的碳和能量的限制是一个普遍规律。

本文回顾了细菌和真菌——土壤中两个主要的微生物界——的碳和能量需求,它们之间对这些和其他资源的竞争机制,竞争机制所导致的生态位分化,以及全球变化对这种竞争的影响。相关研究结果以Mechanisms and implications of bacterial-fungal competition for soil resources为题于2024年5月发表于The ISME Journal。山东农业大学农学院王超群博士和德国哥廷根大学Yakov Kuzyakov为本文作者。
Summary
标准化的微生物利用偏好显示,细菌在吸收简单有机化合物作为底物方面效率高出1.4至5倍,而真菌在利用复杂化合物方面的效率是细菌的1.1至4.1倍。因此,细菌在竞争简单底物时明显优于真菌,而真菌则在利用复杂化合物时占优势。细菌还与真菌竞争在降解复杂底物过程中释放的产物。基于这些细节,我们区分了这两组微生物在土壤中的空间、时间和化学生态位。全球变化的五大主要因素(包括CO2浓度升高、氮沉降、土壤酸化、全球变暖和干旱)将增加这两组微生物之间的竞争。CO2浓度升高、氮沉降和变暖增强了细菌的竞争力,而土壤酸化和干旱则增加了真菌的竞争力。
Carbon and energy demands of bacteria and fungi
细菌和真菌的碳、能量源
所有细菌和真菌的生长、维持和休眠过程都需要消耗能量,其中大多数过程也消耗碳(C)。根据它们的结构复杂性、表面属性及代谢特点,不同微生物群体偏好不同的有机化合物。土壤中的主要碳和能量来源包括根系分泌物、植物凋落物、微生物残体、有机肥料和土壤有机质(SOM)。
植物将光合固定的碳(约10%-50%)分配给根系分泌物,年均约800–4500 kg C·ha⁻¹。根系分泌物主要包括糖、羧酸和氨基酸,因其易溶解,能迅速被细菌和真菌利用。此外,植物凋落物的微生物利用受分解阶段的影响,早期阶段可快速吸收可溶性分子,而后期阶段则需要更多能量和碳来生产外源酶以分解复杂化合物。
微生物残体碳占土壤有机碳的35%-51%,是一个重要的碳和能量来源。微生物残体的降解速度可能比植物残留物中的复杂化合物(如木质素)更快,因为它们的有机化合物较小且具有较高的C:N比。
土壤中主要碳和能量来源的NOSC值按以下顺序增加:脂质、微生物残体、木质素、氨基酸、酚类、植物凋落物、NaOH可萃取SOM、糖类、HCl可萃取SOM和羧酸(图1)。尽管有机化合物中的能量含量随着 NOSC 值的增加而降低,但能量可用性(词汇表框)会增加。总的来说,微生物优先利用具有更高标称氧化态(NOSC)的有机化合物,即使这些化合物的能量含量较低。
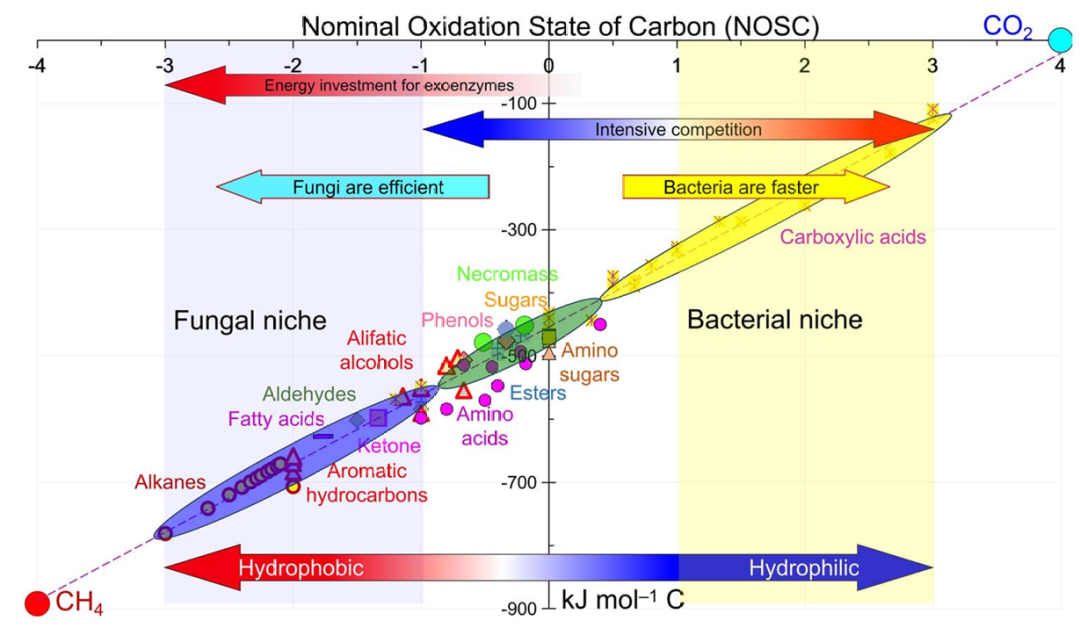
图1. 碳的标称氧化态(NOSC)(横轴)与主要化合物作为土壤中微生物碳和能量来源的能量含量(燃烧焓)(纵轴)的概念表示。
高能势(例如吉布斯自由能 [ΔGo])反映在所有 C 原子的标称氧化态 (NOSC) 中,可以使用以下公式:
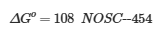
其中 ΔGo 是有机化合物(可能可用于微生物)中的能量含量(焓),NOSC 是这些化合物中所有 C 原子的标称氧化态;后者可以使用以下公式计算:
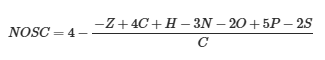
Hydrophobicity of carbon and energy sources
微生物对有机化合物的利用不仅取决于其结构复杂性和能量可用性,还与有机化合物的溶解性密切相关。可溶性有机化合物容易通过溶液扩散,被微生物从更大范围的土壤中捕获,而难溶化合物通常具有较高的分子量,或被吸附在金属氧化物、粘土矿物或有机物上,导致它们较难被微生物利用。
细菌和真菌在对有机化合物的竞争中存在显著差异。细菌通常快速吸收可溶性有机物,而真菌则更擅长利用难降解的有机物(如木质素)。细菌在对水溶性有机物的快速利用上具有优势,因其简单的细胞结构和快速的生长与底物摄取速率(Table 2)。相反,真菌拥有更高的疏水性和更有效的外源酶,能够通过氧化和水解利用疏水性化合物。
水溶性和疏水性有机化合物的差异对微生物的降解过程有重要影响。高疏水性的有机物会减少微生物的分解速度,而水溶性有机物则能更有效地被微生物降解。尽管真菌和细菌都倾向于优先利用水溶性有机化合物,但由于其更高的疏水性,真菌在利用疏水性化合物方面具有一定的竞争优势(Table 2)。
Preferences for carbon and energy sources
为了评估碳和能量来源的偏好,收集了155个数据(详见附表S1),记录了13C标记物质在微生物生物标志物——磷脂脂肪酸(PLFAs)中的掺入情况。设计了以下方程来规范化真菌相对于细菌PLFAs中的13C富集度(13CFungi/13CBacteria),与真菌相对于细菌的生物质碳含量相关:
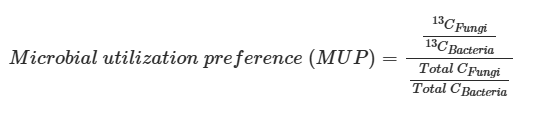
其中13CFungi和13CBacteria分别为真菌和细菌PLFAs中的13C富集度,(Total CFungi)和(Total CBacteria)分别为真菌和细菌PLFAs中的碳含量。较高的MUP值表示真菌相对于细菌在特定底物上有更高的碳吸收。在MUP值大于1时,真菌在该底物的利用上超过细菌,反之亦然。
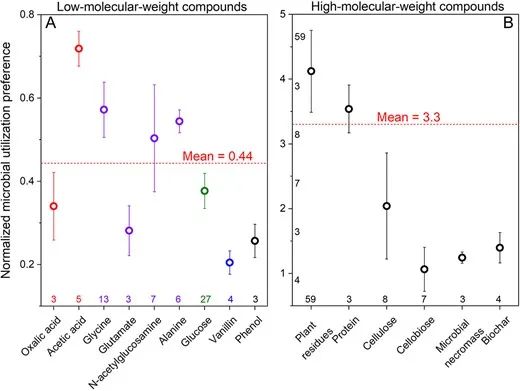
图2 使用各种简单(A)和复杂(B)化合物作为底物,规范化微生物利用偏好(MUP),表示真菌PLFAs相对于细菌PLFAs的掺入情况。红色虚线表示使用简单(A)和复杂(B)化合物作为底物的平均MUP值。MUP值越接近1,表示细菌和真菌在该底物上的竞争力越相似。数据为均值±标准误差。左侧的数字表示研究的数量。请注意,低分子量化合物(左侧)平均MUP偏好为0.44,表示细菌的竞争力是真菌的2.3倍。
MUP值在0.20–0.72之间,表示细菌在小分子(分子量<200 Da)和水溶性化合物的竞争力是比真菌强1.4–5倍(图2A)。相反,复杂底物(例如植物残渣、微生物尸体、蛋白质、纤维素、纤维二糖、生物炭)的MUP值范围为1.1到4.1(图2B)。因此,细菌在简单底物的利用上超过真菌,而在复杂底物方面,真菌具有显著优势。
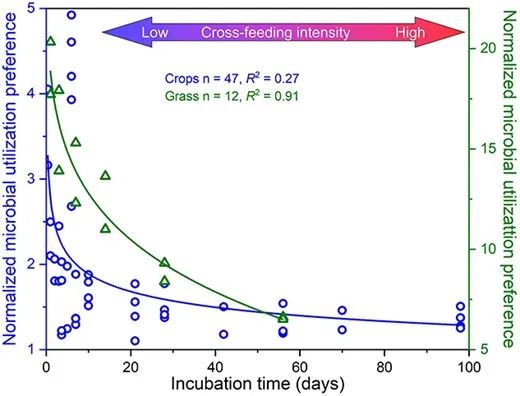
图3 使用作物残体(圆圈)和草类残体(三角形)作为底物时,规范化MUP值的关系,表示真菌PLFAs相对于细菌PLFAs的碳掺入情况与培养时间(即残体分解时间)之间的关系。该关系通过幂回归分析得出(P < .001)。MUP值随着残留物分解的急剧下降,表明细菌与真菌之间的交叉喂养相互作用(大约在10-20天后开始)。
使用植物残体作为底物时,随着分解时间的延长,MUP值逐渐下降(图3),表明真菌在早期分解阶段对植物残渣的竞争力较强。然而,随着时间的推移,细菌通过交叉喂养现象从真菌中直接或间接获得更多产物(图4)。这是因为细菌只能利用植物残渣分解产生的小分子化合物,这些化合物需要时间才能产生。这一点也得到了纤维素和纤维二糖作为底物时较低MUP值的支持(这些底物具有较快的降解速率),与使用分解较慢的蛋白质和植物残渣作为底物时的情况相比(图3)。
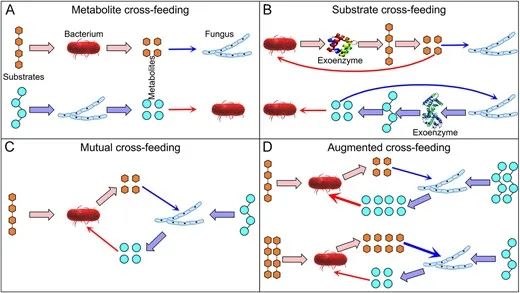
图4 细菌-真菌交叉喂养类型。(A) 代谢物交叉喂养:一种生物体以复杂化合物为食,释放废弃代谢物供另一种生物体利用。(B) 底物交叉喂养:一种生物体分泌外源酶,将复杂化合物分解为简单化合物,供另一种生物体使用。(C) 互惠交叉喂养:代谢物交叉喂养和底物交叉喂养的任意组合,其中细菌和真菌是交叉喂养者。(D) 增强型交叉喂养:互惠交叉喂养的一种子集,其中一种生物体向另一种生物体提供更多的交叉喂养化合物。
交叉喂养通常指一种生物体利用另一种生物体的代谢产物(有机化合物和营养物质)(框1),通常只考虑细胞内代谢。然而,土壤中的交叉喂养通常包括且主要涉及一种微生物群体利用由另一群体通过外源酶产生的外部反应释放的产物。考虑到外源酶反应产物的扩散损失,我们假设土壤中的大部分交叉喂养通过机制2:底物交叉喂养发生(图4B)。尽管交叉喂养相互作用最可能是积极的,但它们会在生物体之间造成竞争。底物交叉喂养是生物体间利用性竞争的基础。尽管细菌和真菌在其他交叉喂养相互作用中不直接竞争产物(图4A、C和D),它们仍会在一些共享资源(例如水、氧气、营养物质)上进行一定的竞争。
Bacterial–fungal competition and niche differentiation
土壤中的碳和能量供应有限是普遍现象,通常限制非丝状细菌、放线菌和真菌的生长。这些限制导致细菌和真菌在有机碳和能量的竞争中持续竞争。
能量竞争比碳的竞争更为激烈。首先,1)微生物可以在细胞内外回收碳,这需要能量投资但不消耗新碳。2)其次,微生物必须投资能量来还原有机和无机化合物,而无需利用碳。3)第三,微生物为维护投入大量能量,各种过程(如细胞分裂、代谢变化、细胞运动、基因表达调控、能量泄漏反应)消耗能量但几乎不消耗碳。例如,细菌在有氧条件下30°C时的维护能量(12 kJ mol−1 C h−1)比真菌(8 kJ mol−1 C h−1)高50%。然而,双方都需要更多能量来相互竞争,从而减少了用于其他功能的能量。
细菌和真菌在竞争易得的碳和能量来源时十分激烈,但它们也进化出竞争和共生的策略来分解难降解的有机化合物。革兰氏阴性细菌通常是快速生长的r-策略者,能够迅速摄取易得的底物,而真菌和革兰氏阳性细菌是生长缓慢的K-策略者,能有效利用难降解的有机化合物。这在根际表现为细菌(尤其是革兰氏阴性菌)的数量远高于真菌,其中根系不断分泌简单底物。
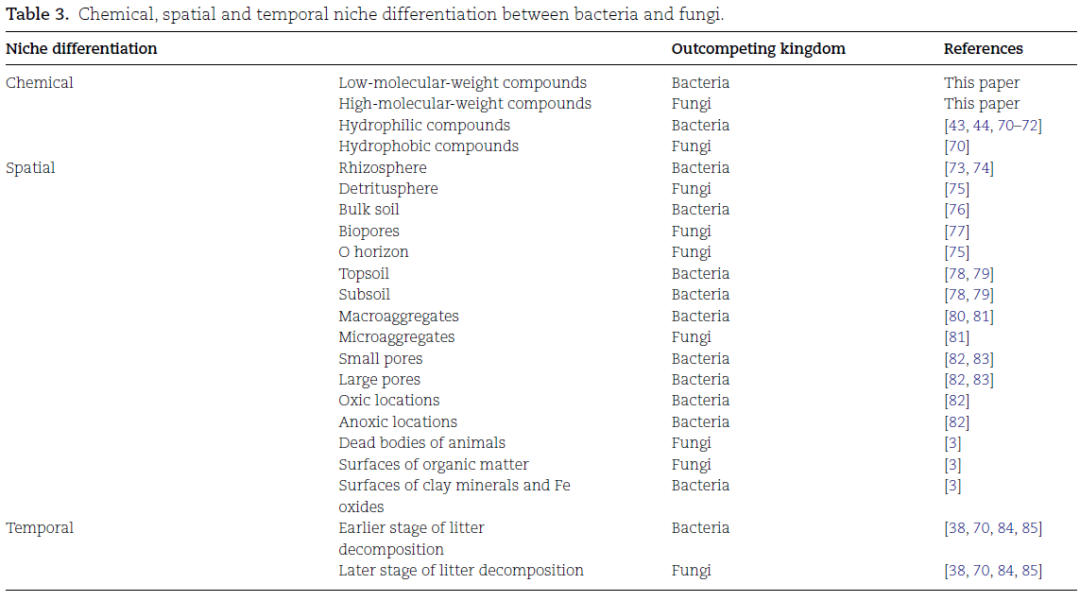
两组生物在土壤中的多个生态位中共存(Table 3),增加了它们对碳和能量的竞争。生态位分化有效地减少了这种竞争。生态位分化的赢家取决于其形态学(如细胞大小、结构、疏水性)、生理学(如生长速率、水和营养需求)和代谢(如酶催化效率、碳和能量利用效率)特性(Table 2)。生态位的物理化学条件(如有机化合物的复杂性、氧浓度、pH值)也起着作用。
Mechanisms of bacterial–fungal competition
竞争类型分为两种:“掠夺性竞争”和“干扰性竞争”(Glossary Box)。细菌和真菌通过这两种类型的竞争以及生态位分化(图5)发展出各种策略,以争夺土壤中的碳和能量资源。
在“掠夺性竞争”中,细菌因多种原因在易得的有机化合物竞争中占优势(图5)。细菌细胞较小,表面积与体积的比率大于真菌细胞,使得细菌能够更快地接触和摄取溶解的有机化合物(表2)。此外,细菌的生长速率远快于真菌(表2),使其能够更快地占领有机物资源。最后,细菌的细胞膜结构比真菌更简单,有利于直接从土壤溶液中摄取小分子化合物。
相反,真菌在竞争复杂有机化合物时占优势(图5),主要因为它们高效地产生几乎所有的外源酶来分解这些结构。真菌能够利用比细菌更丰富的碳和能量来源,因为它们的菌丝增加了栖息地和探索体积(表2),而单细胞细菌则局限于土壤中的小体积。菌丝的物质转运功能和较慢的生长速率(表2)帮助真菌在土壤中维持更稳定的状态,提供了更多的时间来分解复杂化合物。
在“干扰性竞争”中,两组生物通过产生化合物来抑制或杀死对方(图5)。细菌能够产生细菌素(一组小型抗微生物肽或蛋白质)和抗真菌化合物,以减少或甚至停止真菌的生长。反过来,真菌可以产生抗生素(如青霉素、链霉素和四环素,分别由青霉和链霉菌产生)和真菌毒素来限制细菌的生长。两组生物产生的挥发性化合物通过空气孔隙扩散,抑制对方的活性和生长。
细菌和真菌都可以通过形成生物膜来竞争局部的碳和能量资源,排除竞争者(图5)。生物膜通过累积拮抗分子增加竞争力,因为化合物的外向扩散较慢。生物膜还可以保护其中的细菌或真菌免受捕食者的侵害,并促使某些物种向富含碳的区域生长。群体感应是许多物种采取的常见策略,通过调节胞外聚合物物质的产生、表面附着、运动和分散,形成生物膜。通过群体感应,微生物群体可以集体响应竞争,例如通过加大抗微生物化合物的生产并改变其行为,从而获得竞争优势。
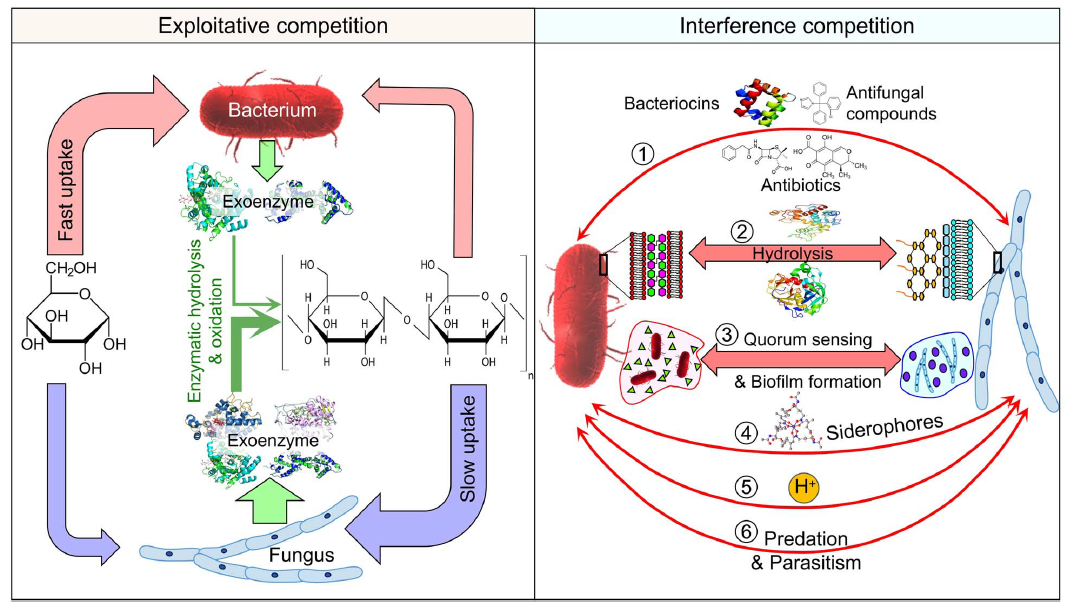
图5 细菌和真菌在利用性(左)和干扰性(右)竞争中争夺碳和能量来源的策略。
在利用性竞争中,细菌快速吸收容易获取的化合物,从而超过真菌;而在复杂化合物上,真菌则通过依赖其多样化的酶和高催化效率来超过细菌。在干扰性竞争中,双方通过以下机制进行竞争(从上到下):(i) 产生各种化合物以抑制或杀死对方,(ii) 分泌特定的水解酶(如几丁质酶、蛋白酶)来分解竞争者的细胞壁或蛋白质,(iii) 产生螯合铁(及其他多价阳离子)的铁载体,以从土壤中夺取铁,从而使需要铁的竞争者得不到足够的铁,(iv) 形成生物膜以增加竞争力,(v) 释放氢离子使土壤酸化,从而为竞争者创造不利条件,(vi) 直接捕食或寄生在竞争者身上。
细菌和真菌都会产生特定的水解酶(如几丁质酶、蛋白酶)来分解竞争对手的细胞壁或蛋白质。两组都产生螯合铁的侧柏素,以从土壤中螯合铁,进而使需要铁来生长和生存的竞争者处于饥饿状态。
微生物还可以通过改变环境的物理化学性质间接抑制对方(图5)。例如,在Collimonas fungivorans和黑曲霉之间的竞争互动中,前者通过产生酸性物质来创造对后者不利的条件。一些代表性物种可能作为捕食者或寄生虫直接竞争生存。真菌食性使得某些细菌可以捕食活的真菌菌丝以获取营养和碳。例如,在侵入土壤中的Absidia菌丝后,Collimonas这一以真菌食性生长而闻名的细菌属,其数量在2周内增加了4倍。
Niche differentiation
生态位分化是减少竞争相互作用的结果和机制。这里区分了化学、空间和时间生态位分化(图6)。
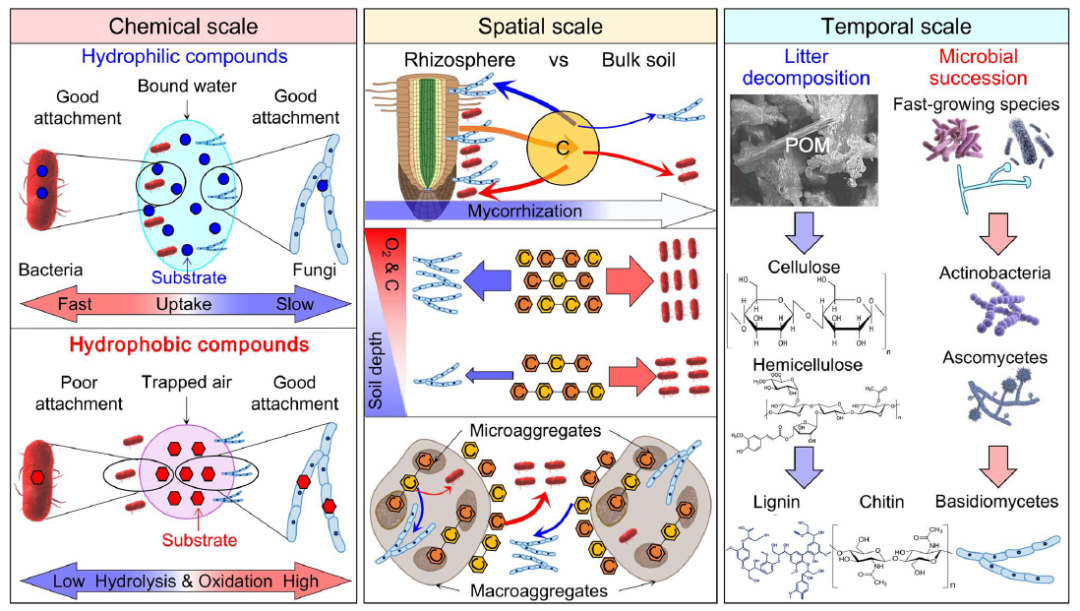
图6 细菌和真菌在化学(左)、空间(中)和时间(右)尺度上的生态位分化。
在化学尺度上,细菌能迅速吸收亲水性有机化合物,从而在与真菌的竞争中占据优势。真菌则通过更有效的外分泌酶和更好的菌丝附着力在疏水性有机化合物上占优。在空间尺度上,(i)真菌在根际比在土壤中更具竞争力,因为根分泌物减少了竞争强度,并促进了菌根真菌的根部定植;(ii)细菌在土壤深度上竞争力更强;(iii)细菌在大团聚体中竞争力更强,而真菌在小团聚体中竞争力更强。在时间尺度上,植物残体分解过程中的微生物演替遵循以下顺序:快速生长的微生物,随后是具有纤维素分解、半纤维素分解和果胶分解能力的真菌和细菌,最后是真菌能够分解木质素和几丁质的过程。
化学生态位分化
细菌与真菌的竞争机制:细菌通过较低的C:N比占优,能更高效地吸收亲水性富氮化合物,如氨基酸和肽类。而真菌则相对依赖其外分泌酶降解疏水性化合物,特别是在植物残体和微生物尸体中含有较多的疏水性有机物。
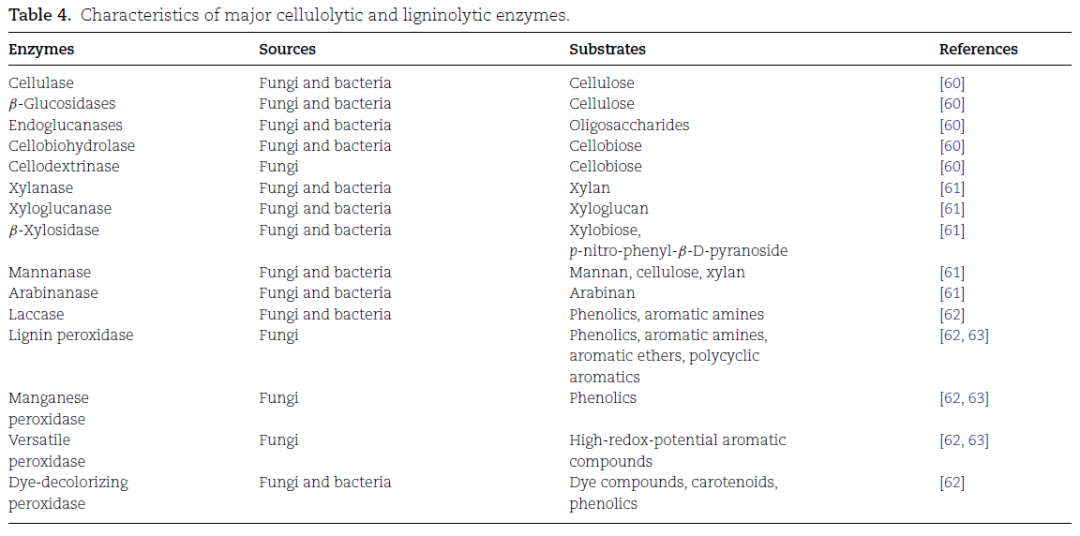
真菌对疏水性化合物的竞争优势:真菌通过分泌氧化酶(如过氧化酶、酚氧化酶)来分解疏水性化合物(表4),这些酶能够将疏水性化合物转化为更亲水的形式,帮助真菌更好地利用这些物质(图6)。尽管某些细菌也能产生氧化酶,但其酶活性和丰度远低于真菌。
外分泌酶的能量投入:微生物在分解疏水性化合物时需要大量投入能量和资源,特别是在外分泌酶的合成上。例如,生产β-葡萄糖苷酶的能量消耗相对较小,但在实际土壤条件下,微生物需要消耗更多的能量来分解复杂的聚合物化合物。
真菌的疏水性适应策略:真菌通过分泌疏水蛋白(如疏水蛋白和表面活性剂)降低表面张力,增强菌丝对疏水表面的附着力,进一步提高其在疏水性有机物上的竞争力(图6)。这些策略使真菌能够有效地在与细菌的竞争中占据优势。
空间生态位分化
有机化合物的空间异质性(如根际和detritusphere碎屑际与土壤、表土与亚表土、大团聚体与小团聚体的差异)导致了细菌和真菌之间的空间生态位分化(表3)。细菌在根际和土壤中竞争有机碳和能量来源时占优势,但在根际,真菌的竞争力更强,因为根分泌物有助于真菌生长(图6)。
根际与残体圈的竞争差异。根际主要选择富营养型细菌,相较于土壤深处,根际中的富营养型细菌比例可高达7倍,这加速了有机碳的消耗,减少了营养较贫乏细菌的竞争力。由于真菌能够直接从根部获取能量,其在根际的竞争力优于在土壤中的表现(图6)。另一方面,真菌依靠高效的外分泌酶系统在复杂有机物丰富的环境中(如碎屑际、生物孔和动物尸体)超越细菌(表3)。
土壤深度对细菌和真菌竞争的影响。随着土壤深度的增加,植物来源的有机碳和土壤有机质(SOM)量减少,导致细菌和真菌的生态位差异。土壤深处的有机质能量含量较低,但其中富含难以降解的疏水性化合物,细菌(尤其是革兰氏阳性菌)在深层土壤中的竞争力更强,而真菌则因需要氧气的生长条件受到限制(图 6;表 3)。
团聚体大小对竞争的影响。不同大小团聚体中的有机碳和能量分布异质性也导致了细菌和真菌的竞争差异。大团聚体中更容易获得有机碳,细菌在其中的竞争力更强;而小团聚体中持久的有机物被矿物质保护,限制了微生物的获取能力,真菌在其中的竞争力相对较高(图6)。这些差异导致大团聚体中细菌主导微生物群落。
时间生态位分化
时间生态位分化体现在细菌在植物凋落物分解初期占主导地位,而真菌则在后期占主导地位。植物凋落物分解过程中的微生物群落变化遵循一个顺序:初期由快速生长的微生物(如细菌、糖类真菌)占据,之后是能降解纤维素、半纤维素和果胶的真菌(如子囊菌)和细菌(如放线菌),最后是能够分解木质素和几丁质的担子菌(图6)。
在分解过程中,细菌和真菌的群落表现出明显的变化。例如,在纤维素添加后的前两周,革兰氏阳性细菌的生物量减少了13%,而革兰氏阴性细菌和真菌的生物量增加了12%-50%。然而,经过两个月后,情况发生了逆转。这些变化表明,随着分解过程中复杂化合物(如木质素、纤维素)的分解,微生物群落的演替过程支持了时间生态位分化的观点。
Glossary Box
抑制共生(Amensalism): A relationship between organisms of two species, in which one is suppressed or destroyed and the other is unaffected. 两种物种之间的关系,其中一种被抑制或摧毁,而另一种不受影响。
共生 (Commensalism):A relationship between organisms of two species in which one organism (commensal) benefits while the other organism (host) of the association is neither benefited nor harmed. 两种物种之间的关系,其中一种有益(共生物),而另一种(宿主)既不受益也不受害。
竞争 (Competition): A relationship between organisms of two species in which both organisms compete for the same resources within an environment at the same time. 两种物种之间的关系,其中两种有机体在同一环境中同时争夺相同的资源。
交叉喂养 (Cross-feeding): An interaction between organisms of two or more species in which metabolic products of one organism are utilized by the other(s). 两种或多种物种之间的相互作用,其中一种有机体的代谢产物被其他有机体所利用。
能量可用性(Energy availability):The ratio of the energy obtained to energy consumed through any activity to the energy that a (micro)organism or community must invest to utilize an organic compound under real soil conditions. 通过任何活动获得的能量与一个(微)生物体或群落必须投资以在实际土壤条件下利用有机化 合物的能量之间的比率。
剥削性竞争(Exploitative competition):Competition in which one population consumes the resources required by another without direct contact between the two populations. 竞争类型,其中一个种群消耗另一个种群所需的资源,而两者之间没有直接接触。
干扰性竞争(Interference competition): Competition in which one population suppresses or stops the growth of another by secreting harmful products. 竞争类型,其中一个种群通过分泌有害物质抑制或停止另一个种群的生长。
微生物残体(Microbial necromass):Microbial residues—the remains of dead microbial cells, cell fragments, cell organelles, and cytoplasm.微生物残余物——死去的微生物细胞、细胞碎片、细胞器和细胞质的遗骸
本文主要总结了“细菌和真菌的碳与能量需求” “碳与能量来源的疏水性” “碳与能量来源的偏好” “细菌与真菌的竞争与生态位分化”四部分,“Global change impacts”和“Coexistence of bacteria and fungi”将在后续推送。
ISME(国际微生物生态学会期刊)(International Society for Microbial Ecology Journal)是由国际微生物生态学会主办、施普林格·自然出版集团出版的微生物生态学领域学术期刊,隶属于Nature子刊体系。其刊载内容涵盖微生物群落、环境互作及生物地球化学循环等研究方向,重点收录在微生物生态系统结构与功能机制中具有突破性进展的研究成果,投稿类型包括原创研究、综述及观点文章等 。
该期刊创刊于2007年,2023年影响因子为10.8,2024年转为完全开放获取模式。作为学会旗舰期刊,与《ISME通讯》形成互补定位:前者聚焦基础性重大发现,后者侧重技术性高质量成果。编委会由国际知名学者组成,中国科研团队曾在该刊发表微生物适应机制、入侵物种生态学等研究成果。

