近日,环境顶刊
《Water Research》
发表了题为
"Convergence and divergence of microbial communities in river- Qinghai lake sediment continuum on Tibetan Plateau"
的研究论文。
文献标题:青藏高原河流-青海湖沉积物连续体中微生物群落的趋同与趋异
文献来源: Li J ,Wang L ,Wu B , et al. Convergence and divergence of microbial communities in river- Qinghai lake sediment continuum on Tibetan Plateau. [J]. Water research, 2025, 282123757.
Doi:10.1016/j.watres.2025.123757
相互连接的河湖系统中的微生物群定义了碳和养分的循环,但微生物群落在从支流过渡到湖泊过程中的组装机制仍有待破译。本研究利用靶向
18S、ITS2
和
16S rDNA
区域的高通量扩增子测序技术,对中国青藏高原咸水湖——青海湖及其相连的上游淡水支流沉积物中的原生生物、真菌和细菌群落进行了研究。研究发现,不同微生物类群的组装机制存在差异:支流中的微型真核生物(原生生物和真菌)群落主要由随机过程塑造(贡献约
85%
),而在湖泊中则转变为环境选择主导(约
55%
)。相反,支流中细菌群落的组装主要由确定性过程主导(约
60%
的环境选择),而在湖泊中则转变为随机过程主导(约
70%
)。尽管存在这些差异,所有类群在多样性和网络复杂性方面都表现出一致的生物地理模式。从支流到湖泊的过渡增强了微生物共现网络的复杂性,但导致了显著的物种损失,
α
多样性降低了
56%–62%
。
β
多样性从支流到河口增加,但在湖泊内部降低。微生物
α
和
β
多样性与沉积物碳氮比呈正相关,与沉积物总碳含量呈负相关。值得注意的是,湖泊沉积物中仅有
1%
至
13%
的微生物类群源自支流,这表明需要进一步的地质调查来探究替代路径。本研究为青藏高原河流-青海湖沉积物连续体中原生生物、真菌和细菌群落的多样性与网络复杂性趋同生物地理模式,以及分异组装机制提供了新的见解。
湖泊作为全球水圈和生态网络的重要组成部分,与上游支流持续相连,在水循环、气候调节和维持全球生态平衡中发挥关键作用。水生沉积物中的细菌、真菌和原生生物等微生物群落具有高度物种多样性,支撑着碳氮循环、有机物降解等生态系统功能,且对环境及人为干扰高度敏感,是衡量环境健康的重要指标。生态位理论和中性理论为微生物群落组装提供了强调确定性过程和随机过程的对立框架,而微生物类群因细胞大小、营养策略等差异组装机制不同,如真菌对营养和地理距离更敏感。生态网络分析显示,拓扑特征可反映群落稳定性,污染物会改变微生物互作模式。青海湖作为青藏高原最大内陆咸水湖,本研究沿其上游支流至湖心纵向采样,假设群落组装从支流的扩散主导转为湖泊的环境选择主导、微生物多样性在过渡中先升后降、网络复杂性随支流汇入增加,以揭示高原河湖微生物分布机制。
1. 实验设计
本研究聚焦青藏高原青海湖及其上游支流,探究沉积物中微生物群落特征与生态机制。
2023
年
7
月,研究人员沿上游至湖心区域布设采样点
(
图
1a、b)
,采集
94
份
0-20cm
浅层沉积物样本,部分样本用于
DNA
提取,部分用于碳、氮、磷等元素组成分析。利用试剂盒提取基因组
DNA
,针对原生生物、真菌和细菌特定基因区域分别进行
PCR
扩增,构建文库后在
Illumina NovaSeq
平台测序。生物信息学分析时,运用
QIIME2
等工具处理原始序列,聚类获得
ASVs
并进行物种注释,同时划分微生物功能群、预测功能谱。数据分析在
R 3.5.1
环境下完成,通过线性回归、
NMDS、PerMANOVA
等方法解析微生物多样性、群落差异及影响因素;借助指示物种分析筛选敏感
ASV
,构建共现网络揭示微生物互作模式;最后使用
βNTI
和
iCAMP
模型评估确定性与随机过程对群落组装的贡献,系统阐明青海湖流域微生物群落的生态过程与机制。
湖泊采样点沉积物中的总碳、总氮含量以及氮磷比均高于上游和河口采样点,而湖泊采样点的总磷含量较低。总碳、总氮含量以及氮磷比随着与湖心地理距离的增加而降低(图 1c、d)。此外,经测序和质量控制后,共获得1,734,162 条 18S序列、1,391,290 条ITS2序列和11,371,928 条16S序列,分别聚类为23,119、12,067和78,854个非单例扩增子序列变体(ASV)。物种鉴定后,数据集中包含6,882个原生生物ASV、10,962 个真菌ASV和77,129 个细菌ASV。稀疏曲线表明,本研究的采样量足以覆盖具有代表性数量的ASV。
原生生物ASV中41%‒46%为上游特有,河口和湖泊特有比例分别为23%‒34%和5%‒9%,三者共有的仅1%‒3%。功能上,原生生物以消费型(52%)为主,其次为光合营养型(12%)和寄生型(6%);真菌群落中腐生型占 65%、寄生型占 26%;细菌则以化能异养(38%)和好氧化能异养(32%)为主。指示物种分析表明,从上游到湖泊的梯度中,多种原生生物、真菌和细菌ASV呈现富集或耗竭(sASV)。河口相比上游,原生生物ASV更多耗竭,而真菌和细菌ASV更易富集;湖泊相比上游和河口,三类微生物ASV均以耗竭为主,且细菌耗竭指数高于真菌和原生生物(图 2a‒c)。这些sASV主要隶属于囊泡虫门、变形菌门、子囊菌门等,其中消费型原生生物、腐生/寄生真菌及参与尿素分解的细菌类群从上游至湖泊显著减少。
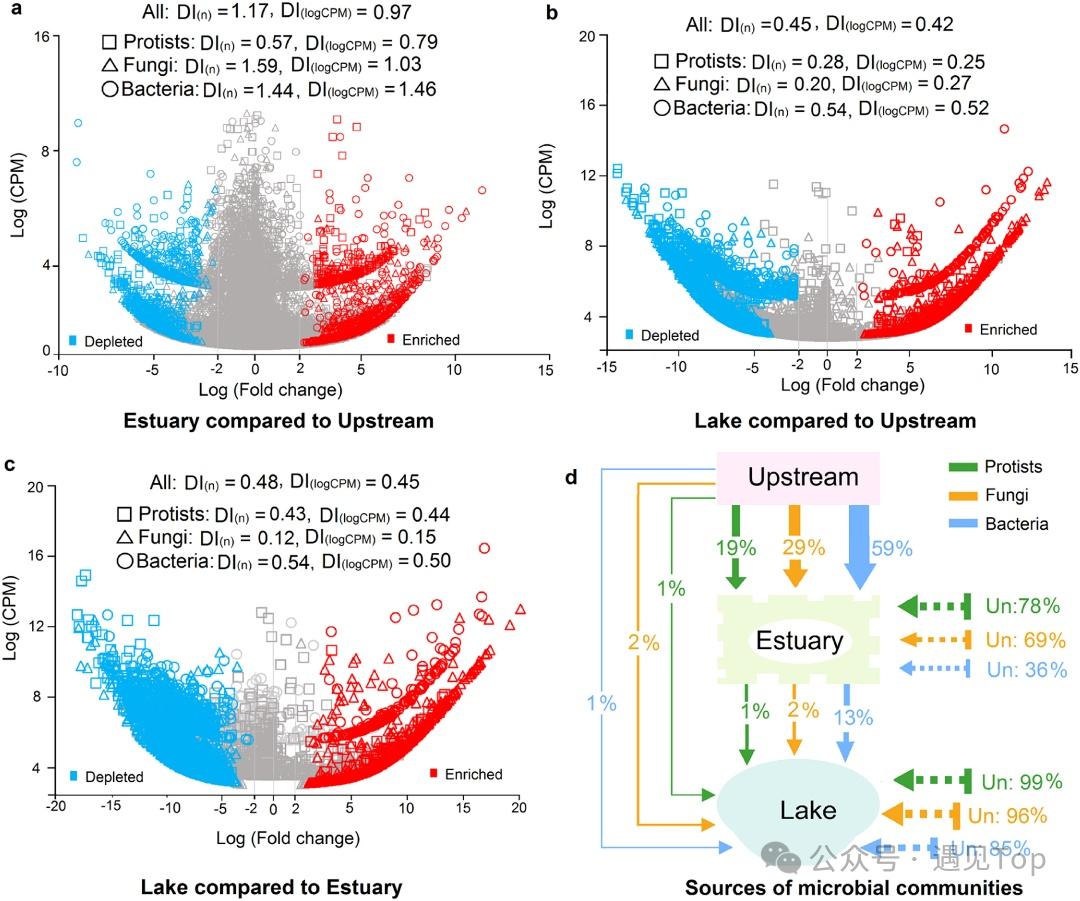
图2. 上游支流、河口和湖泊微生物群落的富集与枯竭及其潜在来源
NMDS 和 PerMANOVA 分析显示,上游、河口和湖泊的微生物群落组成存在显著差异(图 3a)。变异分配分析表明,空间距离对群落变异的解释力高于沉积物碳氮磷含量及比值,而 Mantel 检验证实群落组成与总碳、总氮及碳氮比显著相关,与总磷无关联(图 3b)。
图3. 上游、河口和湖泊生态位中原生生物、真菌和细菌群落组成的非度量多维尺度排序
4. 假设H1-支流和湖泊的主导生态过程,以及湖泊微生物的来源
βNTI模型显示,支流中原生生物与真菌群落组装以随机过程为主(−2<βNTI<2),细菌则以确定性过程为主(|βNTI|≥2),iCAMP模型进一步表明上游和河口的原生生物与真菌受扩散限制和漂变影响,湖泊中则以环境选择为主(图4a,b)。这与原生生物和真菌细胞体积大、扩散能力有限有关,其群落呈现明显的距离衰减模式,而青海湖中的确定性过程可能由盐度、碳氮含量驱动,导致物种耗竭(图2b‒c)。细菌在支流中受确定性过程主导,因体积小易被水流运输,而湖泊中转向随机过程,空间距离解释度最高。SourceTracker分析显示,湖泊中仅1%‒13%的微生物源自支流,支流与湖泊特有类群分别达46%和9%,共享类群仅2%‒5%(图2d),支持“生态岛”理论,暗示地质屏障、群落老化等因素影响微生物分布。
图4. 蓝藻藻华不同阶段真菌-细菌共现模式的演替
5. 假设H2-在支流流向河口和湖泊时原生生物、真菌和细菌多样性的变化
原生生物、真菌和细菌的α多样性呈单峰地理分布,随距湖心距离增加先升后降,真菌对地理距离响应更强(R²=0.1),空间距离是影响α多样性的主要因素。湖泊中三类微生物α多样性较支流降低,可能因上游与河口的物种替换及湖泊高盐度等生境过滤作用(图 3a)。β多样性在河口先升后在湖泊中降低,河口异质性驱动多样性升高,而湖泊的盐度、pH 等物理-生物地球化学过滤导致其下降(图 1c,d)。盐度是淡水向盐湖过渡的关键屏障,β多样性与碳氮比正相关,与总磷无关,且真菌和细菌对总碳响应相反。
6. 假设H3验证:支流至湖泊的微生物网络结构变化
从上游到湖泊的过程中,微生物网络复杂性逐步增强(图 5)。跨界网络显示,原生生物、真菌和细菌间以正相互作用为主,细菌作为关键连接者(图 5a、e),而环境过滤使敏感ASV按生境聚类,如适应湖泊的sASV多聚集在模块4、7、8 中,为 “核心微生物组”(图 5b‒f)。分物种网络表明,从上游到湖泊,三类微生物网络的模块性降低,节点数、边数等增加,网络更复杂均质,与高原盐湖中高盐度放大细菌网络复杂性的结论一致。
图5. 生态位(上游、河口和湖泊)跨界、原生生物、真菌和细菌共生网络结构
在本研究中,探究了青海湖区域从支流到河口及湖泊的过渡梯度上,微生物群落组装机制与生物地理模式。沿河湖连续体,原生生物、真菌和细菌群落呈现出相似的生物地理趋势:与湖泊相比,支流中α和β多样性更高,但网络复杂性更低。然而,群落组装过程在微生物类群和栖息地间存在差异:支流中的原生生物和真菌以随机过程为主导,而细菌群落中确定性过程更为普遍。相反,在湖泊中,原生生物和真菌群落以确定性过程为主,细菌群落则以随机过程更为突出。有趣的是,与预期相反,湖泊沉积物中仅一小部分微生物群源自上游支流,这表明其他来源(可能与微生物群落老化过程相关)需进一步研究。本研究为青海湖河湖连续体中的微观生物地理模式和群落组装机制提供了有价值的见解。
文章链接: Li J ,Wang L ,Wu B , et al. Convergence and divergence of microbial communities in river- Qinghai lake sediment continuum on Tibetan Plateau. [J]. Water research, 2025, 282 123757.

