摘要:石榴对低温敏感,籽粒褐变被认为是低温伤害的典型症状,极大地影响了果实的品质。本文利用广泛靶向的次生代谢组学方法,对正常和褐变的籽粒进行了综合比较。籽粒中共鉴定出399种代谢物,其中褐变籽粒中75个积累上调,14个积累下调。根据《京都基因基因组百科全书》(KEGG)的富集分析,与籽粒相关的植物次生代谢产物生物合成主要涉及类黄酮、黄酮醇和异黄酮,尤其是苯丙素类的生物合成。此外,本研究还进行了籽粒性状与代谢物加权基因共表达网络的相关性分析。总的来说,籽粒褐变主要是由失水、多酚氧化酶(PPO)氧化和水解反应引起的。结果表明,对香豆酸对石榴籽粒褐变的影响较大。这些研究结果将有助于阐明石榴冷藏过程中籽粒褐变的原因,并为石榴采后改善果实品质提供科学依据。
亮点:
1. 首次利用广泛靶向的次生代谢物研究籽粒褐变;
2. PPO的氧化和水解反应是籽粒褐变的主要原因;
3. 对香豆酸对籽粒褐变有较大影响;
4. 褐变籽粒中花色素苷中,只有花青素-3-O-阿拉伯糖苷积累。
原名:Widely targeted secondary metabolomics explored pomegranate aril browning during cold storage
译名:广泛靶向次级代谢组学研究了石榴籽粒冷藏过程中的褐变现象
期刊:Postharvest Biology andTechnology
IF:5.537
发表时间:2022.01
通讯作者:郑先波
通讯作者单位:河南农业大学园艺学院
实验设计

实验结果
1. 正常籽粒(HA)和褐变籽粒(BA)的性状比较
籽粒褐变引起冷藏期间的生理紊乱,影响了石榴果的食用价值。我们比较了突尼斯石榴果HA组和BA组的籽粒性状(表1)。从表中可以看出,BA组的籽粒水分降低(P<0.05),说明冷藏导致重量下降。在籽粒颜色上,BA组的a*和L*值较低,而b*值较高。结合图1A中所示的籽粒外观,正常籽粒呈浅红色,褐变籽粒呈棕色,与表1结果一致。与HA组相比,BA组总黄酮和花青素含量显著增加,而抗坏血酸含量显著降低。两组总酚、SSC、TA含量差异无统计学意义(表1)。此外,MDA积累和EL是评价膜完整性的重要指标。从正常到褐变籽粒,两者均显著上升(表1),表明细胞膜完整性被破坏。
表1 正常籽粒(HA)组和褐变籽粒(BA)组的籽粒性状概况

数据以均数±标准误差表示,差异有统计学意义p < 0.05。
2. 正常籽粒和褐变籽粒酶活性的比较
如表1所示,BA组APX和SOD酶活性低于HA组。BA组POD和PPO酶活性增加,PPO酶活性差异显著,表明PPO对籽粒褐变的影响大于POD。
3. 正常籽粒和褐变籽粒的次级代谢物谱
从图1B的二维主成分散点图中,PC1和PC2两个主成分分别解释了总变量的56.97%和12.04%。3个生物重复的样品沿PC1聚集在同一侧,并能明显区分,说明分析是稳定和可重复的。同时,该图预测代谢物从正常籽粒到褐变籽粒发生了显著变化。基于层次聚类分析(HCA),热图显示了6个样品中所有鉴定的代谢物的积累规律(图1C),层次分类为2个主要分支(一条黄条和一条蓝条),说明两组之间的总代谢物有明显的差异。黄色条来自褐变籽粒,而蓝色条来自正常籽粒。三个重复也分别聚类,这与PCA结果一致。

图1 石榴籽粒次生代谢组学分析
A,在光照条件相同的条件下,拍摄正常籽粒(HA)和褐变籽粒(BA)的表型图。B,次生代谢物的二维PCA散点图。C,HA组和BA组6个样品的次级代谢产物热图。D,石榴籽粒次生代谢物的分类。
UPLC-ESI-MS/MS共鉴定出石榴籽粒中的399个次生代谢物,分为8类:黄酮类148个、酚酸120个、单宁36个、萜类30个、生物碱27个、木脂素和香豆素24个、醌类6个和其他8个代谢物(图1D)。代谢物中黄酮类化合物含量最高,其次为酚酸类、单宁类和萜类。根据VIP ≥1和绝对Log2FC(倍数变化)≥ 1,火山形图被用于筛选差异代谢物。
4. 正常和褐变籽粒中差异代谢物的概况
我们共检测出89种显著调节的代谢产物,其中BA组75种代谢产物增加,14种代谢产物减少(图2A)。从KEGG功能富集图中,89个差异代谢物富集于10条途径,其中2条富集最丰富的途径与代谢途径和次生代谢物的生物合成相关(图2B)。代谢途径主要与酪氨酸代谢和苯丙氨酸代谢有关。次生代谢产物生物合成的相关途径主要与类黄酮、黄酮和黄酮醇生物合成、异喹啉生物碱生物合成、苯丙素生物合成、泛醌和其他萜类醌生物合成有关。热图显示了HA组和BA组差异代谢物相对含量的变化趋势(图2C)。酚酸在积累上调的代谢物中占很大比例,达到32种酚类化合物,如参与苯丙烷生物合成的对香豆酸、咖啡酸和阿魏酸。黄酮类化合物排名第二,在29个差异代谢物中有21个积累上调,其中前3个积累上调代谢物均属于黄酮类化合物,包括2个黄酮醇和1个黄酮类化合物。BA组中只有两种酚酸(5-O-coumaroylshikimicacid O-glucoside和异绿原酸B)减少(图2C)。我们进一步鉴定出7种花青素和8种原花青素。关于花青素,六种花青素衍生物(包括花青素-3-O-阿拉伯糖苷、花青素-3-O-半乳糖苷、花青素-3-O-葡萄糖苷、花青素-3-O-桑布双糖苷、花青素-3,5-O-二葡萄糖苷、和花青素-3-O-(2''-O-葡萄糖基) 葡萄糖苷)和一个天竺葵素(天竺葵素-3-O-葡萄糖苷)被检测到。在籽粒褐变过程中,7种花青素中仅花青素-3-O-阿拉伯糖苷含量较低,8个原花青素中仅原花青素A6含量较高(图2C)。此外,我们在石榴籽粒的黄酮醇中检测到18个槲皮素衍生物,其中4个在褐变籽粒中含量较高(图2C)。

图2 HA和BA组次级代谢产物的变化概况
A,差异代谢物的火山图(火山地图上的每个点代表一个代谢物)。绿点代表积累下调的代谢产物,红点代表积累上调的代谢产物,灰点代表检测到的差异不显著的代谢产物。B,正常籽粒(HA)和褐变籽粒(BA)组差异代谢物的KEGG富集图。这些点代表不同代谢物的数量。从红色到紫色表示p值等级。C,HA和BA组差异代谢物的变化概况。
5. 基于WGCNA的次生代谢产物与籽粒性状的相关性研究
我们利用WGCNA构建共表达网络,确定特征代谢物与籽粒性状之间的关联(图3)。通过WGCNA分析,石榴籽粒中共鉴定出399种代谢物,聚类为两个主要分支,分为4个共表达模块(图3A和B)。此外,86个代谢物位于最大的绿松石模块中,与b*值、AC、MDA含量、PPO和EL呈正相关。相比之下,蓝色和棕色模块中的50个次级代谢产物显示出相反的变化模式(图3B和C)。
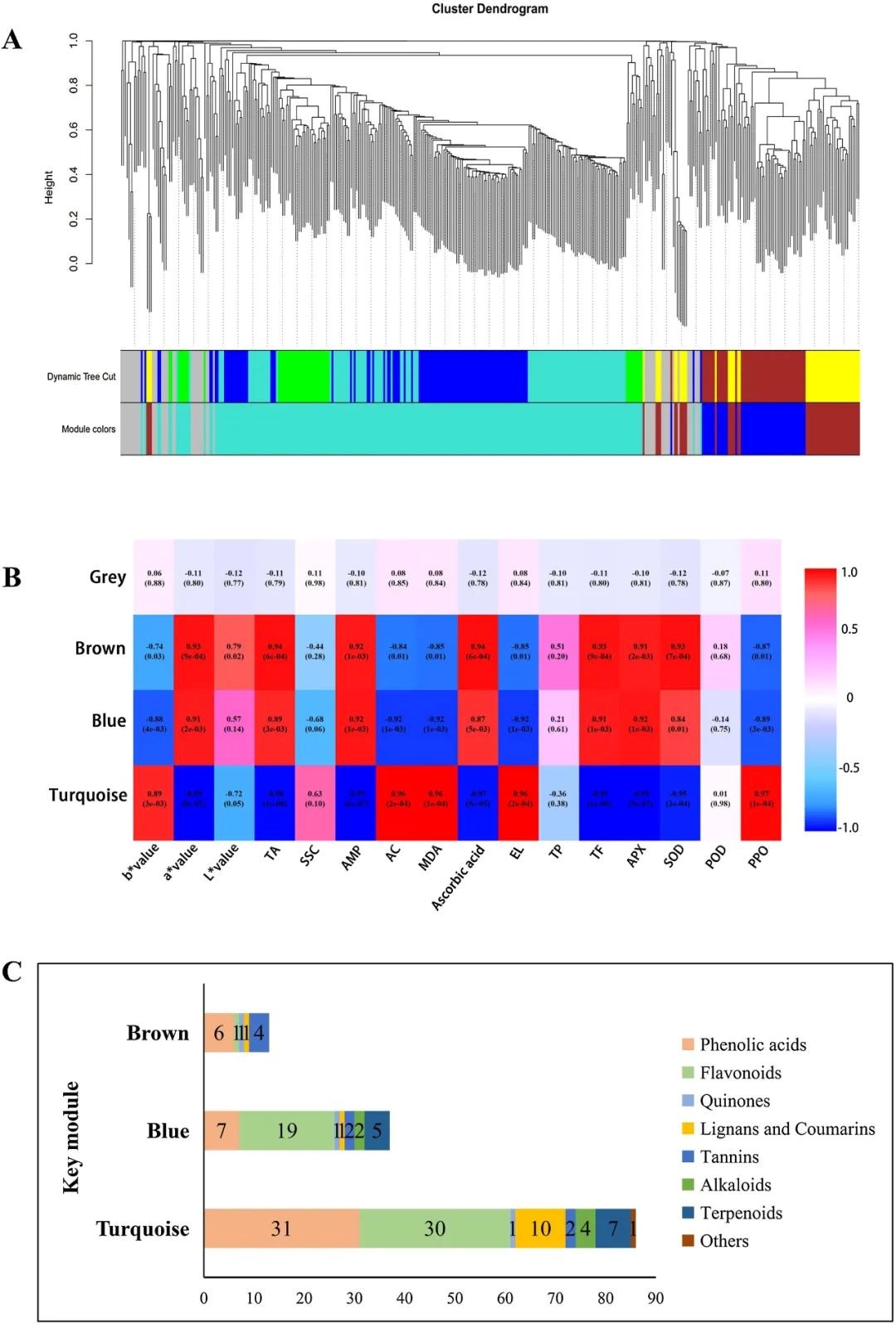
图3 基于WGCNA的代谢产物与生理指标的相关性
A,平均网络邻接度的聚类树形图用于代谢物共表达模块的鉴别。B,模块-性状关系图。C,不同类型的代谢物在关键模块中的分布。注:AMP,籽粒含水率;AC,花色素苷含量;EL,电解质泄漏率;TP,总酚含量;TF,总黄酮含量。
6. 89个差异代谢物与4种相关酶活性的相关性
基于WGCNA,显著差异的代谢物用于评估图4中抗氧化酶活性和氧化酶活性的个体相关性。结果表明,PPO与89种差异代谢物存在密切相关性(与75种代谢物呈正相关,与14种代谢物负相关),而POD与这些代谢物相关性不明显。此外,APX和SOD与这些代谢产物的相关性与PPO相反(图4A)。如图4B所示,丰度代谢物与PPO呈正相关,而积累下调代谢物与APX、SOD呈正相关。

图4 基于WGCNA的显著差异代谢物与酶活性的相关性
A,89个差异代谢物与4种氧化酶和抗氧化酶活性的相关性。B,KEGG代谢途径中差异显著的代谢物与四种相关酶的相关性。未检测到的化合物为黑色,而检测到的化合物为蓝色。在BA组中,积累上调的化合物为红色,积累下调的化合物为绿色。每个代谢物下方的圆点表示其与酶活性(从左到右依次为APX、SOD、PPO和POD)的关系。*和**分别处于p < 0.05和p < 0.01水平。
讨论
石榴中许多次生代谢产物的天然产物和抗氧化活性,如多酚、黄酮类化合物、花青素、抗坏血酸和鞣花酸,已被广泛研究和评估。石榴是一种亚热带和热带果树,极易受冷害和腐烂。采收的石榴果实更容易发生籽粒褐变,并伴有果皮褐变斑,造成营养价值和商业价值的巨大损失,从而大大降低了人们的购买欲望
褐变是一个复杂的过程。当膜被破坏时,酚类物质和酶被释放并与底物接触。总酚含量与酶活性不能单独预测酶促褐变的发生。因此,研究贮藏期籽粒褐变过程中次生代谢产物的积累规律是改善品质性状和阐明籽粒褐变的关键。首先,低温诱导MDA积累和褐变籽粒中较高的电解质泄漏率(表1),然后细胞膜的完整性被破坏,与多酚氧化酶(PPO)的接触加速了籽粒中酚类物质的含量,这与香蕉皮褐变的报道相似。其次,贮藏30d后籽粒水分大量流失是籽粒褐变的原因之一,籽粒出现萎蔫症状(表1和图1A)。此外,Elyatem和Kader(1984)证明,冷害的症状包括石榴果实冷藏期间的重量减轻、棕色变色和籽粒颜色苍白。
本研究中,石榴中399个次生代谢产物主要是酚酸和类黄酮,与Liu等(2021)对石榴果实贮藏过程的研究一致。苯丙素类生物合成中的根皮素是石榴和茶叶中的主要多酚类物质之一。根皮素在褐变籽粒中增多(图2C和4B),这可能是由于根皮苷的水解。类黄酮生物合成中的槲皮素和山柰酚属于黄酮醇苷。石榴籽粒共鉴定出18种槲皮素衍生物和17种山柰酚类化合物,其中,四种槲皮素衍生物和两种山柰酚化合物在褐变籽粒中的相对含量分别高于正常籽粒(图2C和4B),这表明黄酮醇苷发生水解反应。总之,得到的结果可作为直接证据,表明冷藏过程中的水解反应参与了籽粒褐变。

图5 从正常籽粒到褐变籽粒的质量性状和主要显著差异代谢物的特征概述
注:AC,花色素苷含量;AMP,籽粒含水率;EL,电解质泄漏率;TA,可滴定酸;TP,总酚含量;AsA,抗坏血酸含量;TF,总黄酮含量。
PPO氧化反应导致果蔬在贮藏过程中发生质的腐烂是常见的现象。随着贮藏时间的延长,PPO催化能力逐渐增强。在冷藏后褐变籽粒中,总酚类物质含量的降低伴随着PPO活性的增加,并且抗氧化酶SOD和APX呈下降趋势(表1),表明PPO活性是由低温胁迫诱导的,并且酚类底物与PPO发生氧化(消耗)。PPO活性和多酚底物浓度一般与褐变程度有关,可产生棕色素聚合物。然而,我们很难进一步揭示酚类化合物是如何影响石榴籽粒褐变的。目前,我们发现褐变籽粒中酚酸(32)和类黄酮(21)的积累上调(图2和图3),表明这些化合物可能是石榴籽粒褐变的底物或产物。从图2B中可以看出,最富集的代谢途径与酪氨酸代谢、类黄酮和苯丙素生物合成有关。据我们所知,酪氨酸和苯丙氨酸都是对香豆酸生物合成的前体,而且对香豆酸还可以合成咖啡酸和阿魏酸。此外,苯丙素类生物合成过程中对香豆酸的下游产物根皮素和丁素在褐变籽粒中积累上升(图2C和4b)。已有研究报道,在莴苣、苹果、桃、梨中,阿魏酸、咖啡酸、绿原酸等酚酸与PPO催化的褐变反应密切相关。咖啡酸和咖啡酰奎宁酸被认为是洋蓟和莴苣发生褐变反应的底物。褐变籽粒含有相对丰富的对香豆酸、咖啡酸和阿魏酸,以及两个对香豆酸衍生物(对香豆酸甲酯和对香豆酸-4-O-葡萄糖苷)和一个咖啡基奎宁酸(3,4,5-三咖啡基奎宁酸)(图2C和4b),说明这些酚酸可能被PPO催化,导致籽粒褐变。原花青素是缩合单宁,是苯丙素和类黄酮途径的最终产物。本研究在石榴籽粒中共检测到8种原花青素,由2种A型、4种B型和2种C型组成。褐变籽粒中原花青素A6的丰度较高(图2C),这是由于B型原花青素通过PPO氧化反应转化为A型原花青素,而该反应受温度的显著影响。此外,大黄素和芦荟大黄素的8-O-糖苷属蒽醌类化合物,首次在石榴籽粒中被发现。大黄素和芦荟大黄素分别呈现浅橙色和深棕色。其中大黄素-8-O-葡萄糖苷含量在褐变籽粒中增加(图2C),说明氧化反应发生在正常转变为褐变籽粒中。结果表明,PPO氧化反应是石榴籽粒褐变的主要原因。在冷藏过程中,对籽粒褐变的关键氧化酶是PPO而不是POD(图3B和图4),这也得到了Maghoumi等人的支持。我们的研究结果也为苯丙素类生物合成是石榴籽粒褐变的重要途径提供了新的观点,尤其是通过抑制酪氨酸酶活性控制黑色素形成的对香豆酸可能对石榴假种皮褐变有较大的影响,对香豆酸通过抑制酪氨酸酶活性来控制黑色素的形成,对石榴籽粒褐变有很大的影响。
花青素是一种水溶性类黄酮化合物,是石榴的主要色素,是石榴籽粒的代表性颜色。正常籽粒从一个红褐色的小点开始,然后蔓延到整个籽粒,形成褐变(图1A),另一方面,这种颜色变化对应于花青素的积累,b*值的增加,以及L*和a*值的减少(表1),低温花青素积累增多也支持这一推论。早些时候的报道发现,石榴贮藏期间的籽粒大部分归因于红色花色素苷的快速降解。在籽粒中,花色素苷主要由花青素和飞燕草苷组成。此外,颜色还取决于总花色素苷和不同花色素苷积累的差异。结果表明,褐变籽粒冷藏期间,色素呈现出不同的变化规律。首次在籽粒中检测到的花青素-3-O-阿拉伯糖苷在褐变籽粒中相对含量呈下降趋势(图2C),而其他花青素和天竺葵素苷的变化不显著,说明石榴褐变的籽粒颜色与7个花青素衍生物中的花青素-3-O-阿拉伯糖苷有很大关系。总的来说,花色素苷的组成,而不是总花色素苷的含量,可能是影响健康和褐变籽粒颜色差异的决定因素。
在褐变籽粒中积累的其他次级代谢产物中,黄酮类生物合成中的二氢杨梅素具有很强的抗氧化活性,还具有许多药理功能。二氢杨梅素在褐变籽粒中的相对含量低于正常籽粒(图2C),且与APX和SOD呈正相关(图4A),表明二氢杨梅素通过降低抗褐变作用参与籽粒褐变。
结论
本研究利用广泛靶向次级代谢组为研究籽粒褐变提供了首次探索。结果表明,籽粒中含有399种代谢产物,其中,褐变籽粒中含有75个积累上调代谢产物和14个积累下调代谢产物。结果表明,籽粒褐变的主要原因是PPO氧化、失水和水解反应。从图5可以看出,低温诱导的膜受损和籽粒枯萎为酚类化合物和PPO的混合提供了潜在的条件。次生代谢产物的差异明显证明籽粒褐变的发生。在各种次生代谢产物中,苯丙素类生物合成中的对香豆酸对于籽粒褐变至关重要,对香豆酸既能抑制黑色素,又是咖啡酸、阿魏酸、紫铆素和根皮素生物合成的前体。此外,在7种花色素苷衍生物中,只有花青素-3-O-阿拉伯糖苷在褐变籽粒中上调,表明花色素苷组成,而非其总含量,影响籽粒褐变和颜色。这些发现有助于解释石榴籽粒褐变的原因,并为石榴贮藏中的关键代谢产物的应用提供有价值的信息。
